| |
|
| |
|
|
|
|
| |
|
| |
|
|
|
|
| |
|
| |
|
|
|
 |
 |
|
INTERACTION
OF COLLOIDAL TRANSPORT AND PREFERENTIAL FLOW
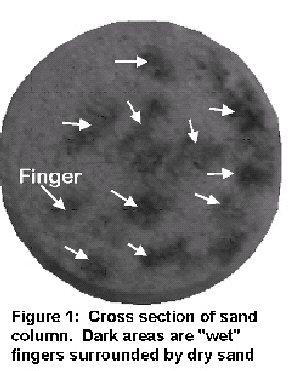
Preferential flow is known to contribute to groundwater pollution of toxic chemicals when a small percentage of the amount applied at the surface reaches the groundwater. The movement of non-adsorbed chemicals through preferential paths is better understood than adsorbed chemicals. Adsorbed chemicals may adhere to colloidal material dissolved in the water or adsorb to the soil matrix directly. This topic is concerned with the interaction of colloidal transport and preferential flow paths. Roots can be one of the causes of the preferential flow paths and the origin of the colloids. With the assumption that the whole soil matrix is participating in the transport process, it is generally considered that colloidal transport in the vadose zone is limited because of film straining and sorption to the air-water (AW) interface. However, under preferential flow conditions, only a small portion of the matrix takes part in the flow process with near saturated flow conditions facilitating transport of colloids. Despite that the breakthrough of a non adsorbed tracer and colloids in the preferential flow paths occur at the same time, our pore scale visualizations reveal that a major portion of the colloids are captured near the air water solid interface of pendular rings. The interaction of preferential flow and colloid transport is well illustrated by an experiment of Darnault et al. (2002) in which the transport of Cryptosporidium parvum oocysts and a tracer were added to 50 cm long columns with coarse sandy soil with grain diameters equivalent to 0.85 - 1.70 mm. Cryptosporidium parvum oocysts are colloids and range in size from 4 - 6 micro meter. The experiments consisted of establishing steady state water flow conditions with an application rate of 2 cm/h. After steady state was established over a billion Cryptosporidium parvum oocysts and the chloride tracer were added as a pulse and water application was continued at the same rate. Both the oocysts and the chloride in drainage water were measured. The columns were segmented at the end of the experiment and the flow pattern and oocysts distribution were observed. In Figure 1 a cross section of the columns is depicted with distinct fingers and dry soil in between. Previous experiments in our laboratory have shown that the moisture content in the fingertip is near saturation. The moisture content becomes more unsaturated farther above the tip. The number of fingers depends on the flow rate (Selker et al., 1996). At a low flow rate fewer fingers will form. This is shown in Figure 1 where there are a relatively large number of fingers because of the high flow rate of 2 cm/hr. This is important for colloid transport since film flow, as postulated in the conceptual colloid transport models, will not occur even at lower flow rates under preferential flow conditions.
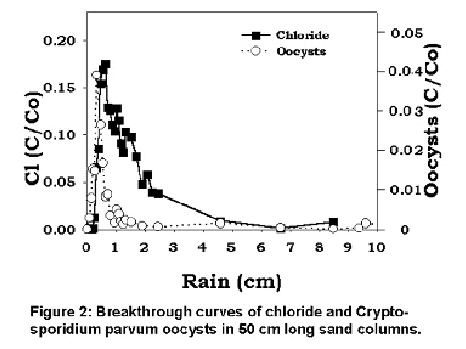
Figure 2 is an example of one of the oocysts and chloride breakthrough curves. The average moisture content in the columns was less than 0.05 cm3 in most of the column. However, because of the high moisture content in the fingers, Cryptosporidium moved readily downwards and appeared at the same time in the drainage water as the chloride. The tail of the Cryptosporidium breakthrough was much smaller than for chloride, indicating that the Cryptosporidium was retained in the column and taken out of solution by some kind of mass transfer reaction.
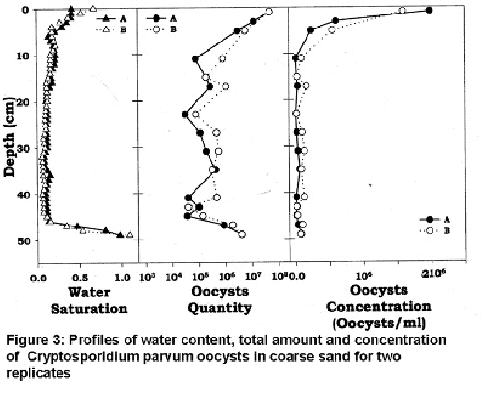
The retention of colloids is confirmed by analyzing the amount of oocysts remaining in the soil (Figure 3). Most of the oocysts are retained near the surface but otherwise the colloids were pretty well uniformly distributed throughout the column with slightly higher concentrations where the moisture contents are greater. Since film flow did not occur in our experiment, film straining as envisioned by Wan and Tokunaga (1997) cannot explain the retention of colloids in the soil profile. It is of interest to examine by pore scale visualizations if other mechanisms can explain the colloid retention. The mechanism mentioned in the literature consists of sorption at the air-water (AW) interface (Wan and Wilson, 1994) or at the solid-water (SW) interface by capture in small pore restrictions and/or by deposition. The sorption of colloids at the air-water interface is the common explanation given for colloid retention in colloid breakthrough studies by Schafer et al. (1998), Jewett et al. (1999), and Lenhart and Saiers (2002).
References: Darnault, C.J.G., P. Garnier, Y-J. Kim, K.L. Overson, T.S.Steenhuis, J.-Y. Parlange, M.Jenkins, W.C. Ghiorse, and P.C. Baveye. 2002. Preferential transport of Cryptosporidium parvum oocysts in variable-saturated subsurface environments. Water Environment Research. In Press. Jewett, D.G., B.E. Logan, R.G. Arnold and R.C. Bales (1999). Transport of Pseudomonas fluorescens strain P17 through quartz sand columns as a function of water content. Journal of Contaminant Hydrology 36: 73-89. Schafer, A., P. Ustohal, H. Harms, F. Stauffer, T. Dracos and A.J.B. Zehnder (1998). Transport of bacteria in unsaturated porous media. Journal of Contaminant Hydrology 33:149-169. Selker, J.S., T.S. Steenhuis, and J.-Y. Parlange. 1996. An Engineering Approach to Fingered Vadose Pollutant Transport. In: Fingered Flow in Unsaturated Soil: From Nature to Model, T.S. Steenhuis, C.J. Ritsema, and L.W. Dekker, Eds. Special Issue of Geoderma 70:197-206. Wan, J.M. and T.K. Tokunaga (1997). "Film straining of colloids in unsaturated porous media: Conceptual model and experimental testing." Environmental Science & Technology 31: 2413-2420. Wan, J.M. and J.L. Wilson (1994). Visualization of the role of the gas-water interface on the fate and transport of colloids in porous media. Water Resources Research 30: 11-23.
|
||||||||||||||||||||||